3.1. Anoxia Experiments
Anoxia was used to chemically isolate the wall from the protoplasm of the sporangiophores of
P. blakesleeanus and to reveal the wall chemistry and chemorheology that produces irreversible wall extension and elongation growth. The sporangiophores of
P. blakesleeanus strictly require oxygen for growth and development [
17,
19]. The relationship between elongation growth and the metabolism of the sporangiophores was studied by using anoxia to inhibit oxidative phosphorylation [
17]. Bergman
et al. [
17] report, “Many fungi can grow anaerobically for long periods of time, but
Phycomyces sporangiophores do not have this capacity. When oxygen is removed from sporangiophores by placing them in nitrogen, streaming and growth rapidly stop.” Studies conducted in our laboratory by Morgan A. Scott (Master’s report, see Acknowledgments) show that streaming inside the sporangiophore stops within a minute;
t = 39.5 ± 2.4 (SE) s,
n = 10. The finding that the turgor pressure continually decreases after the initiation of anoxia is consistent with the results of earlier studies that anoxia terminates the protoplast’s metabolism.
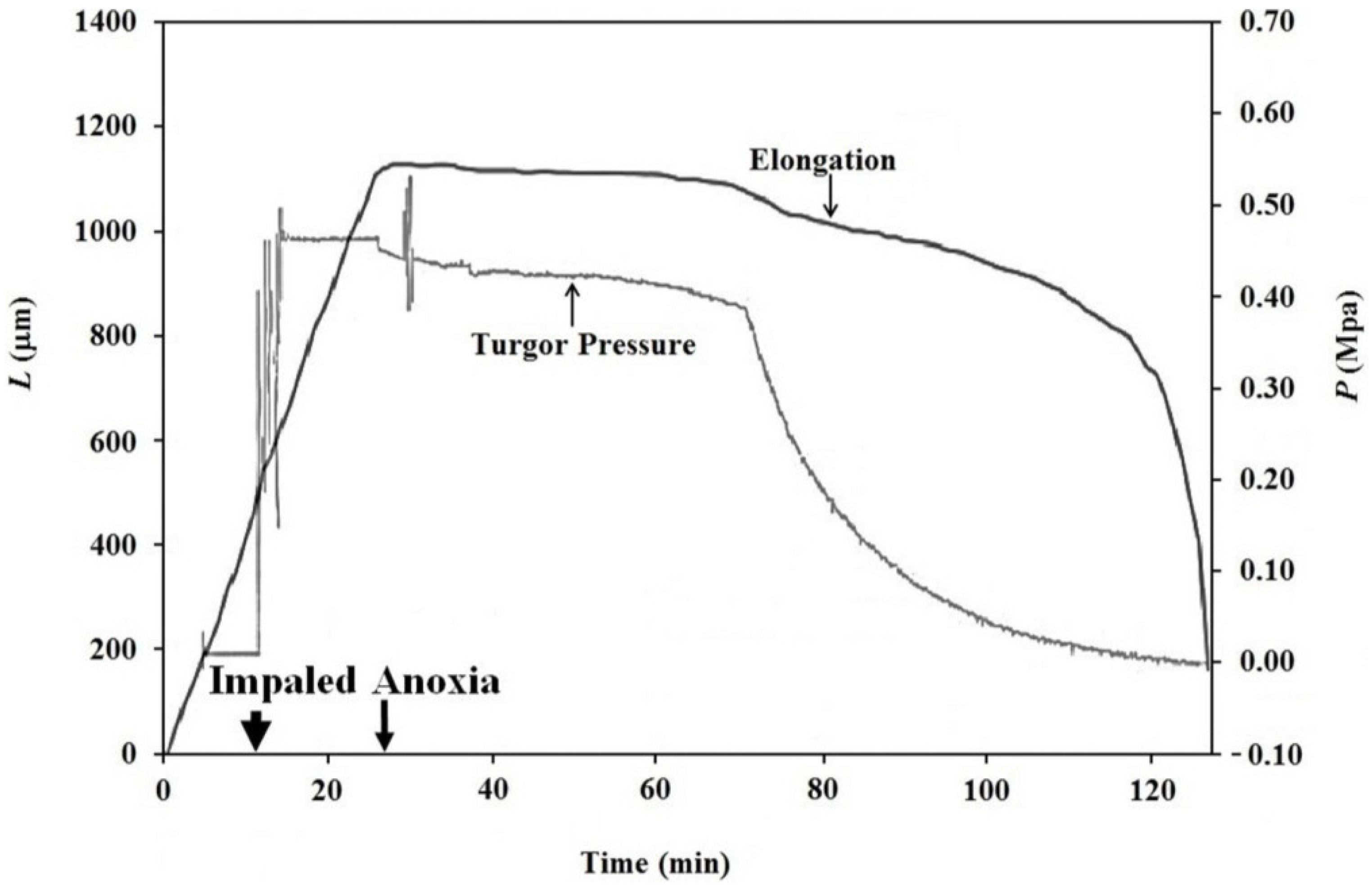
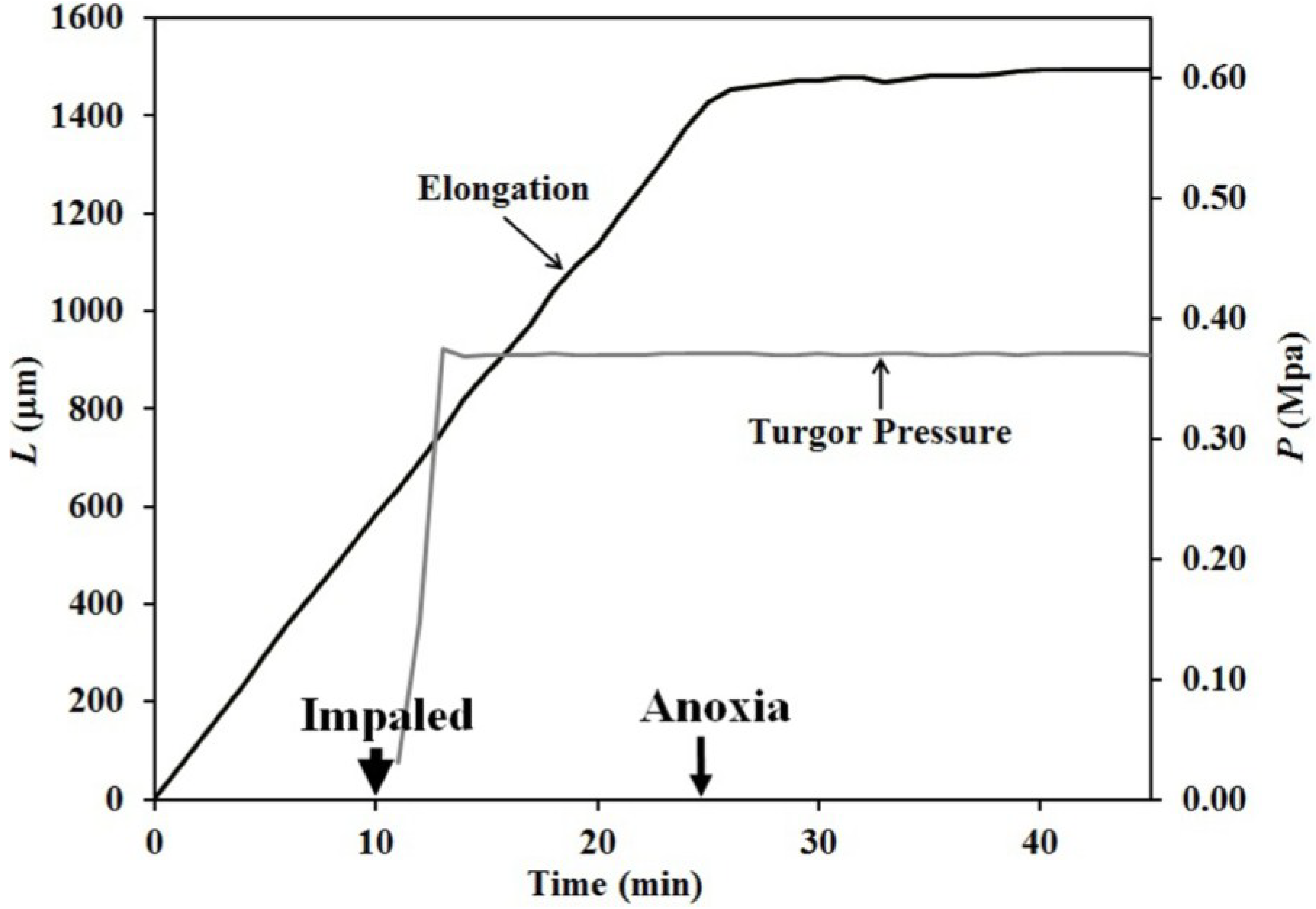
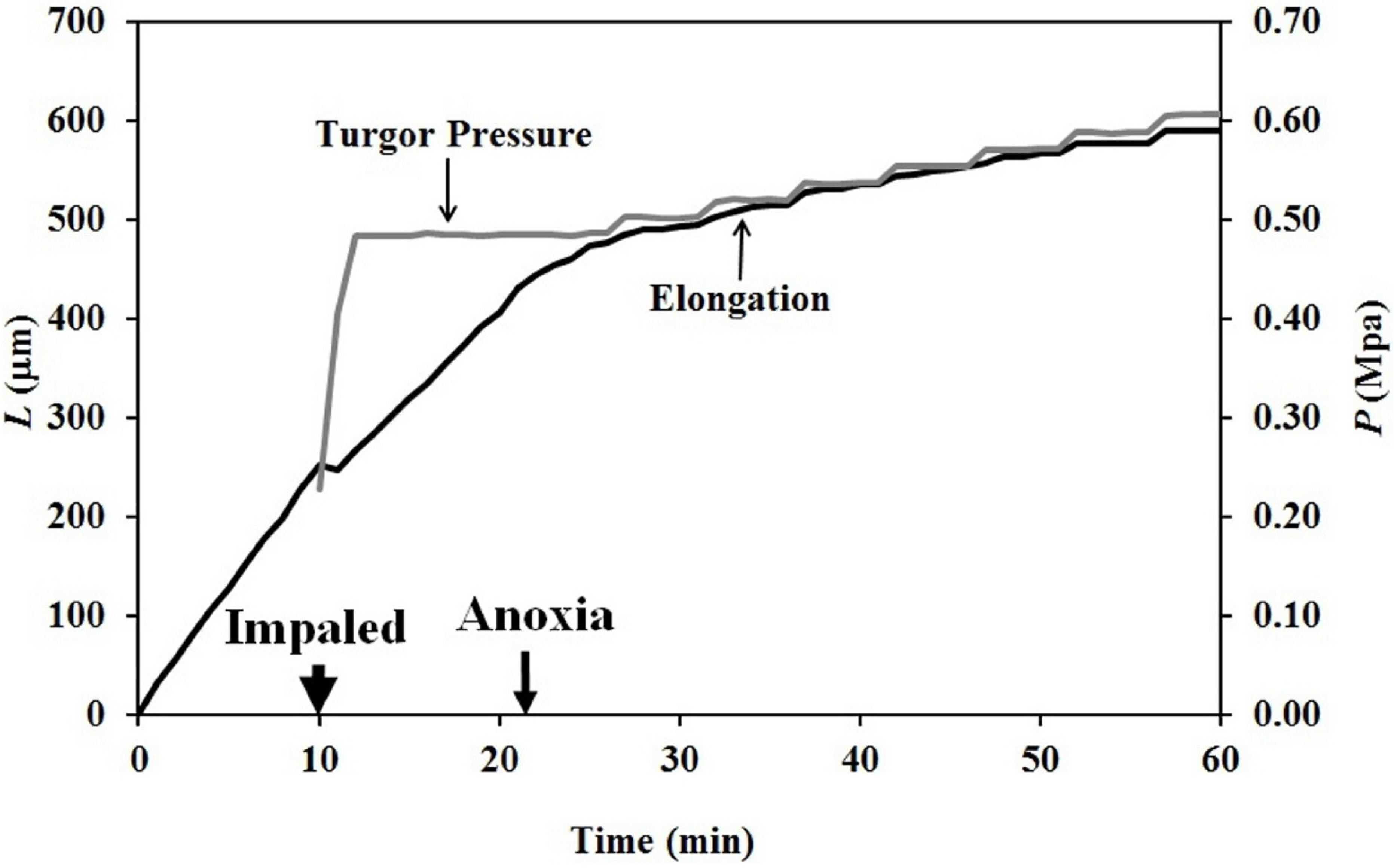
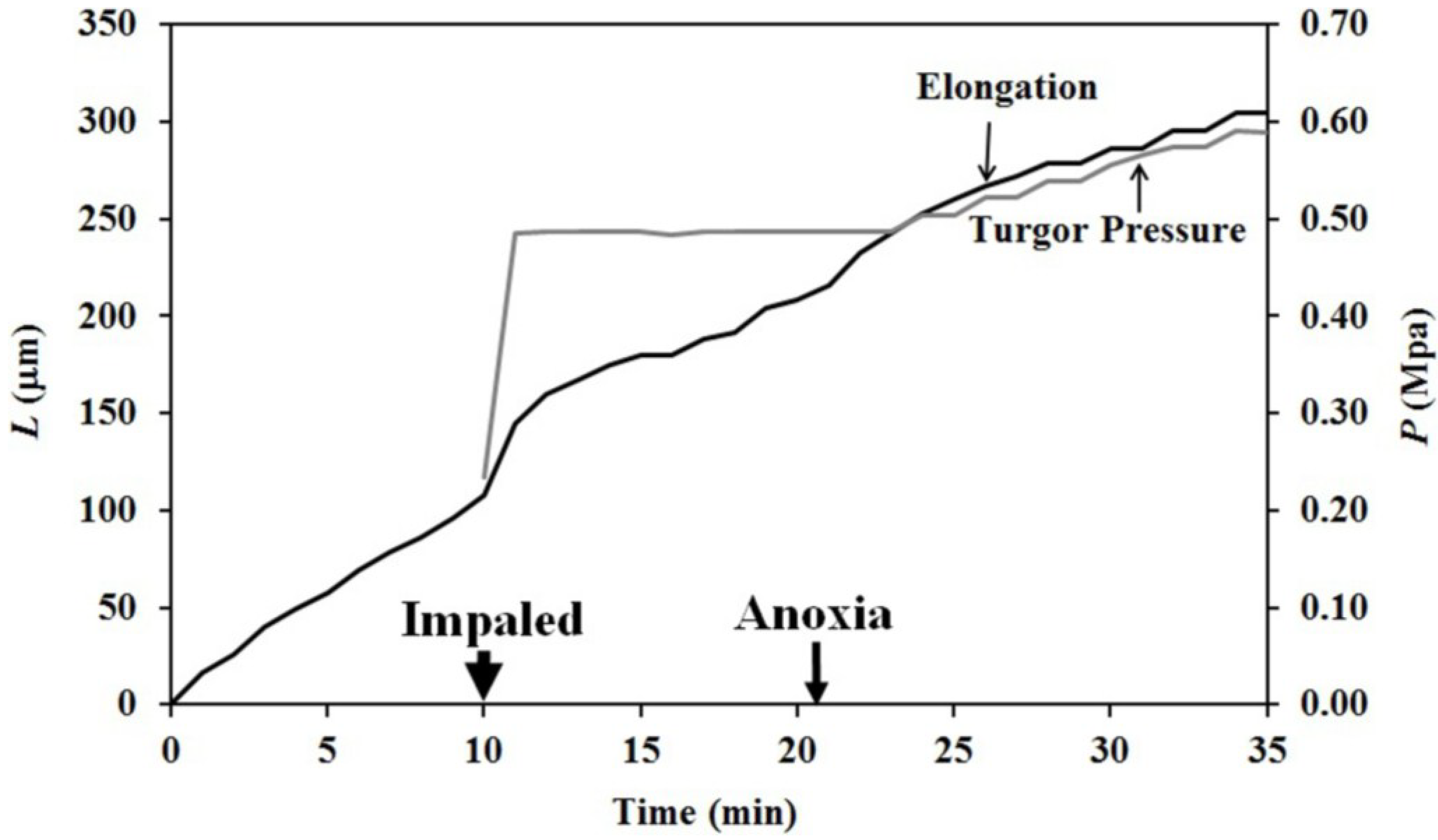
The finding that elongation growth continues during anoxia for ten minutes or longer when the turgor pressure is held constant with the use of a pressure probe demonstrates that a wall chemistry continues the chemorheological process to produce irreversible wall deformation during anoxia. The wall deformation is irreversible, because the elongation growth occurs when the turgor pressure is constant (
Figure 3,
Figure 4 and
Figure 5). Inspection of the previously-established augmented growth equation, Equation (2), can show this. The augmented growth equation can be modified to describe the elongation growth of cells with a growth zone, such as the sporangiophores of
P. blakesleeanus; Equation (3) [
20].
where
L is the length of the sporangiophore, d
L/d
t is the elongation rate,
mg is the longitudinal irreversible wall extensibility of the growth zone,
P is the turgor pressure,
PC is the critical turgor pressure and
Lg and
Ls are the lengths of the growth zone and stalk, respectively.
Eg and
Es are the longitudinal components of the volumetric elastic modulus within the growth zone and non-growing stalk, respectively. The term,
mg (
P − PC), represents the longitudinal irreversible deformation rate of the wall in the growth zone; the term, (
Lg/
Eg) d
P/d
t, represents the longitudinal elastic deformation rate of the wall in the growth zone; and the term, (
Ls/
Es) d
P/d
t, represents the longitudinal elastic deformation rate of the wall in the non-growing stalk.
It is noted in Equation (3) that when the turgor pressure is constant (
P = constant, then d
P/d
t = 0), the elongation rate is only a function of the irreversible longitudinal deformation rate of the wall in the growth zone; Equation (4). Equation 4 is essentially the same equation previously derived by Lockhart [
21]. Thus, the elongation rate that occurs when
P = constant (for 15 min after the initiation of anoxia in
Figure 3, between 2–5 min after the first five turgor pressure step-ups in
Figure 4 and after the first two turgor pressure step-ups in
Figure 5) represents the irreversible wall deformation rate.
The irreversible wall deformation and elongation growth that occur during anoxia are direct evidence of a wall-loosening chemistry or chemorheological process. In the sporangiophores, the wall chemistry and chemorheological process continue for a limited duration during anoxia, typically 10–20 min. Two explanations for the limited duration of elongation growth and associated wall-loosening chemistry during anoxia are considered. The first explanation is that irreversible wall deformation continues during the wall hardening process that begins at the initiation of anoxia. The wall hardening process converts an irreversible extensible wall to a reversible extensible wall by making bonds between microfibrils and other wall polymers. It is predicted that the irreversible wall deformation stops when the wall hardening process is complete. Second, wall building polymers and other relevant wall materials extruded into the periplasm and inner wall surface before the initiation of anoxia, are used to continue the wall-loosening chemistry and maintain the irreversible wall extensibility. After these polymers and materials are depleted, then the wall hardening process begins and continues until completion.
The first explanation predicts that the duration of elongation growth during anoxia for Stage I and Stage IV sporangiophores are approximately the same, because it is expected that the wall hardening process for both stages is similar and the rate of hardening is approximately the same. The second explanation predicts that the duration of elongation growth for Stage IV sporangiophores is larger than those of Stage I sporangiophores, because the elongation growth rates of Stage IV sporangiophores (34.1 ± 3.1 (SE) μm·min
−1,
n = 20) are significantly larger than those of Stage I sporangiophores (7.1 ± 0.7 (SE) μm·min
−1,
n = 17) [
7]. The larger elongation growth rates of Stage IV sporangiophores require a faster delivery rate of wall-building polymers and relevant wall-building materials to the periplasm and inner wall surface compared to Stage I sporangiophores. It is reasonable to expect that the amount of wall-building polymers and relevant wall-building materials within the periplasm and inner wall surface at the time anoxia is initiated (and the wall is chemically isolated) is larger for Stage IV sporangiophores compared to Stage I sporangiophores. If the amount of wall building substrates for the wall chemistry is larger for Stage IV, it is predicted that the duration of the wall chemistry, wall loosening, irreversible deformation and elongation growth will be longer for Stage IV compared to Stage I. The second prediction is consistent with the results presented in
Table 1.
3.2. Frozen-Thawed Sporangiophores
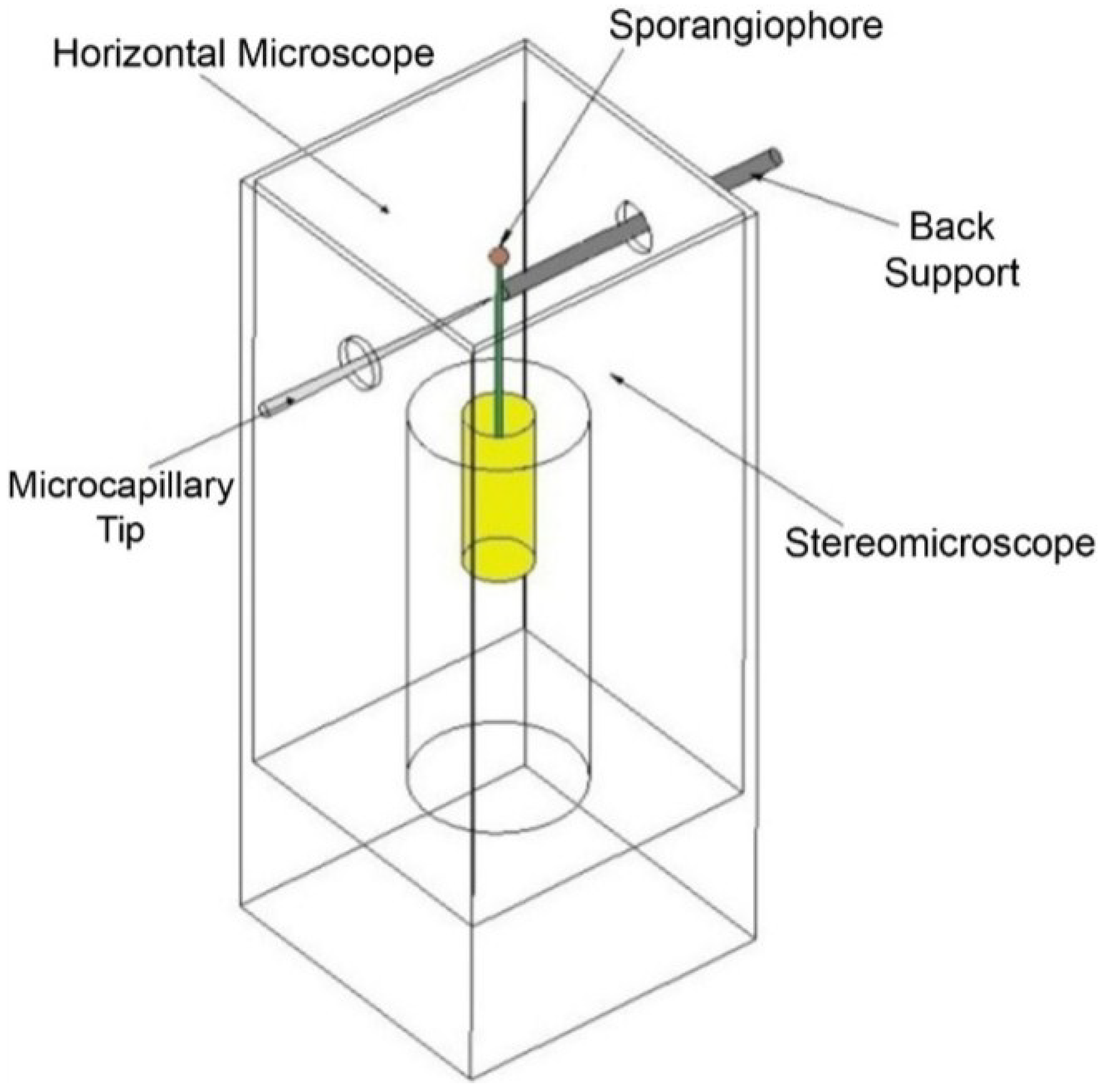
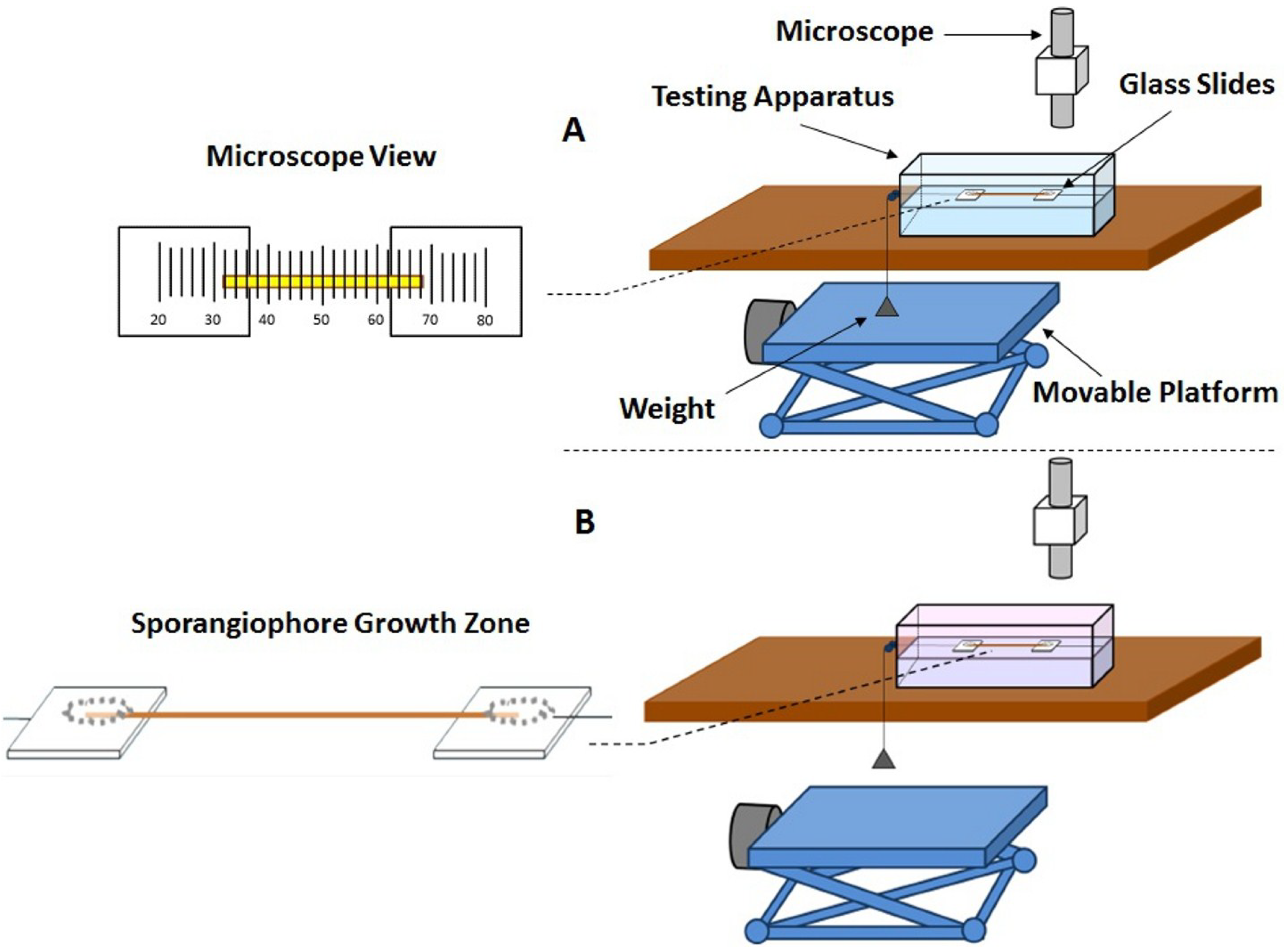
Extension experiments were conducted on frozen-thawed sporangiophore walls to obtain insight into the wall loosening chemistry. The constitutive equation for a Maxwell–Bingham viscoelastic model, Equation (1), can be modified to assist in the interpretation of the results. Equation (5) is derived from Equation (1) and describes the longitudinal deformation (extension) rate of the frozen-thawed wall section after a tensile load, or longitudinal stress (
σ), is applied.
where
L is the length of the frozen-thawed wall section, d
L/d
t is the longitudinal deformation (extension) rate,
mf is the longitudinal irreversible wall extensibility of the frozen-thawed growth zone wall section,
σ is the longitudinal stress,
σC is the critical longitudinal stress,
Lo is the initial length and
Ef is the longitudinal elastic modulus. The term,
mf (
σ – σC), represents the longitudinal irreversible deformation rate of the wall in the growth zone, and the term, (
Lo/
Ef) d
σ/d
t, represents the longitudinal elastic deformation rate of the wall in the growth zone.
Equation (5) is similar to an equation previously used by Takahashi
et al. [
22] to analyze the relationship between the extension behavior of frozen-thawed tissue sections of cucumber hypocotyls and expansin. Here, in Equation (5), we explicitly recognize that
mf (longitudinal irreversible wall extensibility that results from unidirectional stress) is not equal to
ϕ or
mg (irreversible wall extensibility or the longitudinal component of the irreversible wall extensibility, respectively, that results from multidirectional stresses produced by turgor pressure
in vivo), and
Ef (the longitudinal elastic modulus that results from unidirectional stress) is not equal to
ε (the volumetric elastic modulus that results from multidirectional stresses produced by turgor pressure
in vivo).
Constant tension-extension experiments were conducted using a tensile force of 1.24 gf, or 0.012 N (
F = mass × gravitational acceleration = (1.24 × 10
−3 kg) (9.81 m·s
−2) = 0.012 N) that was applied to the frozen-thawed walls. The applied longitudinal wall stress can be estimated using an average sporangiophore diameter of
D = 150 µm and a wall thickness of
τ = 0.6 µm;
σ =
F/π
Dτ ≅ 42 MPa. Immediately after applying the longitudinal wall stress (tensile force of 0.012 N), initial longitudinal wall deformations (extensions) ranged from 1–2 mm. The initial wall extension in response to the application of the wall stress can be determined using Equation (5), where d
σ/d
t is finite and relatively large (typically, the wall stress of 42 MPa is applied within a few seconds). It is apparent that the initial longitudinal extensions are predominately elastic; d
L/d
t = (
Lo/
Ef) d
σ/d
t. After the initial longitudinal extension, the longitudinal stress and length of the frozen-thawed wall section remained constant. Because
σ is constant in Equation (5) (
i.e., d
σ/d
t = 0), subsequent extensions that occur after decreasing the pH of the bathing solution to 4.6 represent irreversible deformations and creep, Equation (6).
The applied longitudinal wall stress,
σ ≅ 42 MPa, is larger than the estimated longitudinal stress produced by the turgor pressure
in vivo. Previously, the pressure probe was used to determine the average turgor pressure (
P = 0.32 ± 0.01 (SE) MPa,
n = 20) and average critical turgor pressure (
PC = 0.26 ± 0.01 (SE) MPa,
n = 20) ([
7] and the references within). The longitudinal stress produced by turgor pressure can be estimated using the equation,
σ =
PR/2
τ, where
P is the turgor pressure,
R is the radius (75 µm) and
τ is the wall thickness (0.6 µm). The longitudinal wall stress produced by the average turgor pressure (
P = 0.32 MPa) is estimated to be,
σ = 20 MPa, which exceeds the estimated critical wall stress produced by the average critical turgor pressure (
PC = 0.26 MPa),
σC ≅ 16.3 MPa.
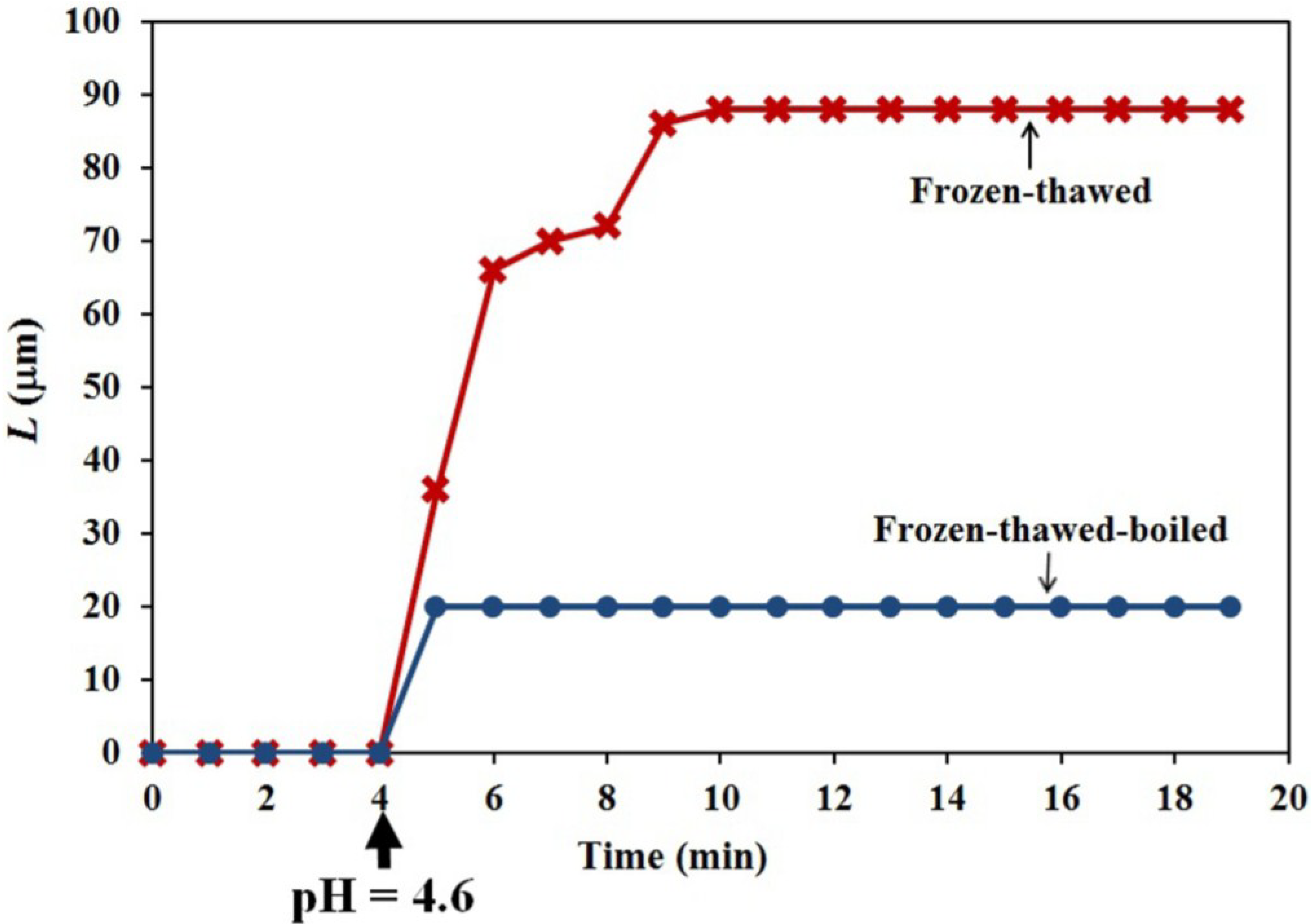
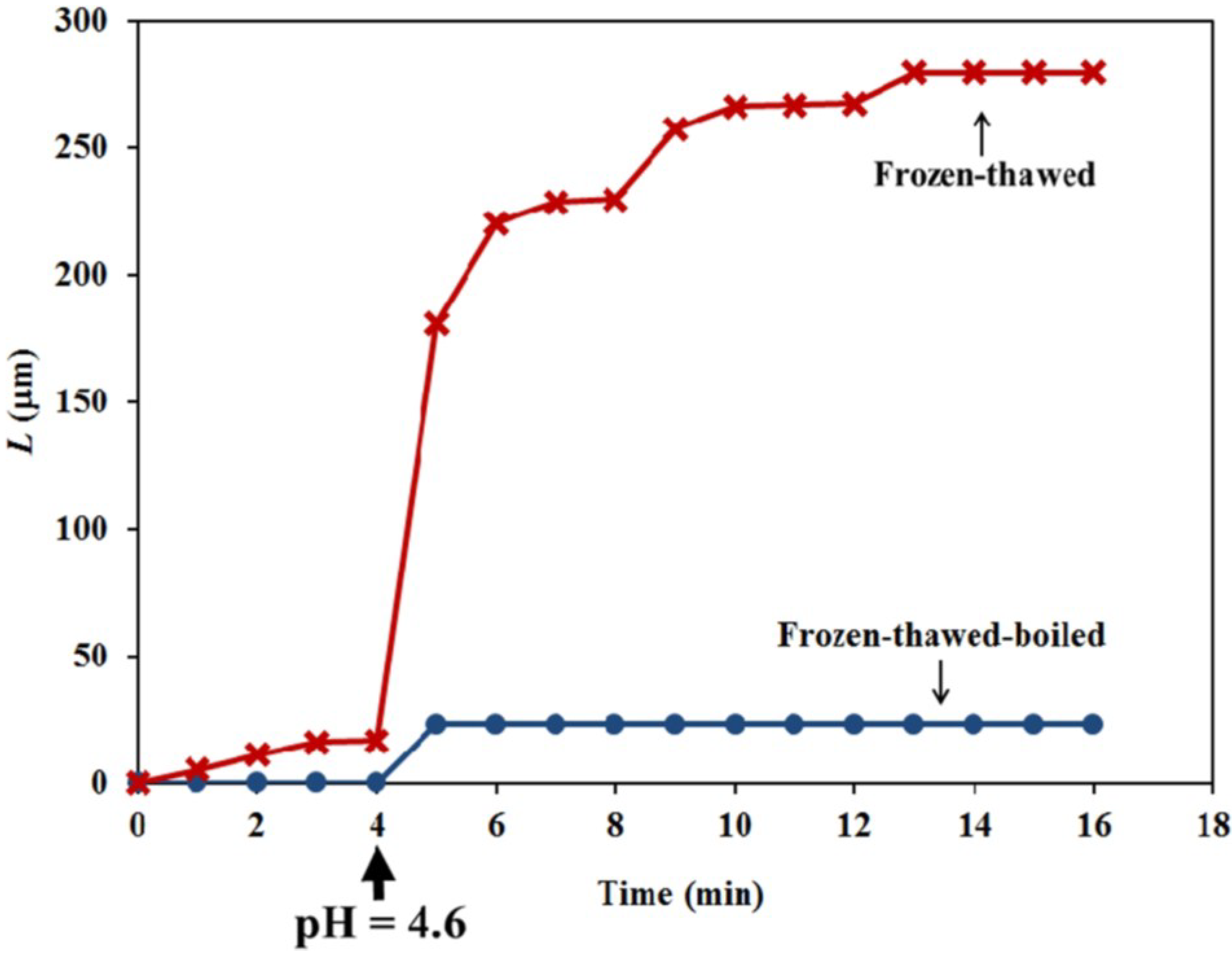
The objective of the constant-tension extension experiments conducted on frozen-thawed sporangiophore walls is to learn if lowering the pH of the bathing solution will elicit irreversible extension and creep, similar to what was found in higher plant cells. The results shown in
Figure 6 and
Figure 7 (red curve with X data points and labeled “Frozen-thawed”) demonstrate that lowering the pH of the bathing solution from 7.0 to 4.6 elicits a chemorheological process, irreversible wall extension (see Equation (6)) and wall creep. In contrast, if the wall sections were immersed in boiling water for 15 s before the constant tension-extension experiments, they did exhibit a nearly instantaneous irreversible deformation, but did not exhibit creep, after the pH of the bathing solution was reduced to 4.6.
Constant-tension extension experiments were previously conducted on frozen-thawed sections of growing cucumber (
Cucumis sativus L.) hypocotyls and oat (
Avena sativa L.) coleoptiles immersed in a bathing solution [
10,
11]. When the pH of the bathing solution was reduced to 4.5, the frozen-thawed walls begin to extend, and the extension continued for hours afterwards, thus exhibiting creep. The experimental results are consistent with the “acid growth” hypothesis [
9], which states that plant cells excrete protons onto the inner surface of the cell wall to decrease the pH and activate an “unknown” wall loosening process. When the frozen-thawed cucumber and oat walls were boiled in water for 15 s before the extension experiment, the walls did not exhibit creep when the pH of the bathing solution was lowered to 4.5. These and other experimental results suggested that protein activity may mediate wall loosening at low pH. Subsequently, wall-loosening proteins were isolated that are not enzymes, expansins [
1]. Expansins are hypothesized to catalyze wall stress relaxation and irreversible wall deformation directly by disrupting hydrogen bonds between the cellulose microfibrils and hemicellulose [
23]. Expansins have been found in a variety of plants and in bacteria and fungi [
1,
12]. Crude extracts and purified expansins have been shown to mediate “acid-induced” extension of boiled isolated primary walls of a variety of plants [
1]. Wall loosening by expansins is consistent with the acid growth hypothesis. Expansins are thought to be strong candidates for primary wall-loosening agents for cells in the organs of higher plants (e.g., stems, roots and leaves). However, endogenous wall enzymes, such as xyloglucan endotransglycosylase/hydrolase and endo-β-1,4-glucanases, are also hypothesized to play an important role in expansive growth of higher plants [
13]. Importantly, a considerable amount of experimental evidence indicates that pectin plays a major role in the expansive growth of pollen tubes ([
14] and the references within) and algae ([
15] and the references within). Furthermore, pectin may play a role in the expansive growth of higher plant cells [
3,
8]. Interestingly, a small amount of pectin is found in the sporangiophore’s cell wall [
24], and one wonders if pectin plays a role in the expansive growth of sporangiophores and fungal cells in general.
The general structure of the sporangiophore’s wall can be described as chitin and β-glucans microfibrils embedded in an amorphous matrix composed predominately of chitosan, glycoproteins and lipids [
16,
18], with a small amount of pectin [
24]. The wall polymers are linked together by covalent bonds, hydrogen bonds, hydrophobic interaction and ionic associations [
16]. The microfibrils are extruded onto the inner wall surface by chitin synthases embedded in the plasma membrane [
16]. Chitin synthases and other enzymes and wall polymers are transported to the plasma membrane in vesicles (chitosomes) and delivered to the periplasm via exocytosis [
16]. Importantly, fibrillogenesis and microfibril networks have been shown to occur
in vitro by incubating purified chitin synthases with substrate (UDP-GLcNAc) and activators [
25,
26]. Furthermore, more recently, the genome of
P. blakesleeanus was sequenced, and genes involved in sensory growth responses and cell wall components are being characterized (
http://genome.jgi-psf.org/Phybl2/Phybl2.home.html).
Generally, the microfibrils can be cross-linked directly (by hydrogen bonds, hydrophobic interaction and ionic associations) and by matrix polymers (chitosan, glycoproteins, and pectin). We hypothesize that wall chemistry and molecular agents that effectively break and make load-bearing cross-links between microfibrils produce chemorheology that regulates wall mechanical properties and wall deformation behavior. Breaking the cross-links directly, or by disconnecting the microfibrils from the matrix network and/or each other, will allow the microfibrils to separate and/or slide passed each other when the wall is stressed by turgor pressure in vivo and by an applied stress in extension experiments. In turn, this chemorheological process can produce a controlled polymer creep, often referred to as “wall loosening”. Load-bearing cross-links that are not broken or that were broken and reconnected (after slippage and/or separation of microfibrils) and subsequently become load-bearing produce the elastic behavior of the wall. The wall loosening will initiate wall stress relaxation and turgor pressure relaxation that, in turn, produces water uptake, irreversible wall deformation and an increase in cell volume.
The extension curves obtained from frozen-thawed and frozen-thawed-boiled sporangiophore walls after the pH is lowered to 4.6 are qualitatively similar to those obtained for cucumber (
Cucumis sativus L.) hypocotyls and oat (
Avena sativa L.) coleoptiles [
10,
11]. The experimental results from the frozen-thawed sporangiophore walls are consistent with the “acid growth” hypothesis [
9]. An important difference is that the duration of the creep for the sporangiophores’ walls is limited to less than ten minutes, compared to hours for cucumber hypocotyls and oat coleoptiles. The finding that boiling the sporangiophore walls for 15 s eliminates creep may suggest that protein activity mediates the creep response, but other explanations must also be considered until more experiments are conducted.
The finding that the duration of the creep for the sporangiophores’ walls is limited to less than ten minutes, compared to hours for cucumber hypocotyls and oat coleoptiles, is interesting and probably significant. This finding, together with the fact that the molecular polymers, structure and endogenous enzymes in the sporangiophore’s wall are different from those in plant and algal cell walls, may indicate that the molecular wall loosening mechanism is different for fungal cells. It may be significant that the duration of elongation growth during anoxia and the duration of creep in frozen-thawed walls sections are both on the order of ten minutes. Interestingly, the duration of creep produced by decreasing the pH is approximately the same as the duration of the light growth response and avoidance growth response (transient increases in elongation growth rate) exhibited by the sporangiophores [
7,
17,
18]. A mechanism is suggested by these findings in which proton effluxes mediate sensory growth responses of the sporangiophores. One may speculate that the sporangiophore’s protoplast excretes protons at an increased rate onto the inner surface of the growth zone wall to elicit a transient increase in irreversible wall extensibility [
27,
28] that produces a transient increase in elongation growth rate, without increasing the turgor pressure [
29]; see Equation (4). The phototropic response (growing towards a light source) and avoidance response (growing away from solid objects) could be produced by spatially increasing the proton efflux on the distal side (phototropic response) and proximal side (avoidance response) of the sporangiophore’s growth zone. Future research will include adding wall building substrates, activators and endogenous wall enzymes to the bathing solution during constant-tension extension experiments to learn if they will increase the duration of creep in the frozen-thawed sporangiophore wall.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



