Download as docx, pdf, or txt
You might also like
- E01V Assignment 8Document8 pagesE01V Assignment 8kish0622Noch keine Bewertungen
- Let's Read and Find Out Science BooksDocument6 pagesLet's Read and Find Out Science BooksSharon Orzechowski Weil0% (1)
- H06D Assignment 8Document4 pagesH06D Assignment 8Good ChannelNoch keine Bewertungen
- Cell DivisionDocument21 pagesCell DivisionOlogunja PaulNoch keine Bewertungen
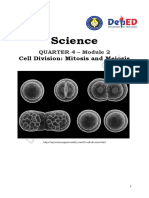
- Science8 Q4 M2Document6 pagesScience8 Q4 M2Arabella BaldonadoNoch keine Bewertungen
- Bio 001 Cell DivisionDocument8 pagesBio 001 Cell DivisionKnick KnackNoch keine Bewertungen
- MeiosisDocument3 pagesMeiosisJaja Jr. Evasco EmbarqueNoch keine Bewertungen
- Cellular Eproduction and GeneticsDocument2 pagesCellular Eproduction and GeneticsmikaelaquisidoNoch keine Bewertungen
- Meiosis: Week 4 DiscussionDocument9 pagesMeiosis: Week 4 DiscussionJacob CarlNoch keine Bewertungen
- Module 2 Meiosis and MitosisDocument88 pagesModule 2 Meiosis and MitosisSevdred CadelinaNoch keine Bewertungen
- MeiosisDocument2 pagesMeiosisZendrix PauloNoch keine Bewertungen
- 6b SamiDocument21 pages6b Samidanyaalahmad7Noch keine Bewertungen
- Meiosis I & 2Document17 pagesMeiosis I & 2Mar Delos ReyesNoch keine Bewertungen
- Cell Division LessonDocument9 pagesCell Division LessonGerry100% (1)
- Cell Cycle and Cell DivisionsDocument19 pagesCell Cycle and Cell DivisionsForkensteinNoch keine Bewertungen
- Cell Division - Mitosis and MeiosisDocument48 pagesCell Division - Mitosis and MeiosisJean CabigaoNoch keine Bewertungen
- Lesson 1Document4 pagesLesson 1catalancsj10Noch keine Bewertungen
- GeneticsDocument4 pagesGeneticsrizamaevillanueva79Noch keine Bewertungen
- Reviewer in General Biology 1Document7 pagesReviewer in General Biology 1gericj7dsNoch keine Bewertungen
- (Biology Form 4) Stage in MeiosisDocument6 pages(Biology Form 4) Stage in MeiosisFuzieanna BakhtiarNoch keine Bewertungen
- Mitosis DefinitionDocument19 pagesMitosis DefinitionMar Delos ReyesNoch keine Bewertungen
- The Cell CycleDocument72 pagesThe Cell CycleJerry Jeroum RegudoNoch keine Bewertungen
- Script (2 Copies)Document3 pagesScript (2 Copies)Zendrix PauloNoch keine Bewertungen
- VC1 - Cell Cycle and Cell DivisionDocument17 pagesVC1 - Cell Cycle and Cell DivisionAnubhab DasNoch keine Bewertungen
- Prelim Task 4Document1 pagePrelim Task 4Kagami TaigaNoch keine Bewertungen
- Heredity and Variation NotesDocument19 pagesHeredity and Variation NotesAmelia DavisNoch keine Bewertungen
- SZL B206 Principles of GeneticsDocument28 pagesSZL B206 Principles of Geneticsawuors249Noch keine Bewertungen
- Module 2 Cell Cycle and Cell DivisionDocument9 pagesModule 2 Cell Cycle and Cell DivisionBalolot RalphNoch keine Bewertungen
- Cell CycleDocument12 pagesCell CycleVega, Charles Gabriel G.Noch keine Bewertungen
- Cellular Reproduction and GeneticsDocument47 pagesCellular Reproduction and GeneticsJean Marie Macadaeg OrdinarioNoch keine Bewertungen
- Mitosis MeiosisDocument8 pagesMitosis MeiosisSyndy Mae Wasing DosogNoch keine Bewertungen
- Cell Division: Dr. P. Pokorski University of Michigan - Dearborn Department of BiologyDocument52 pagesCell Division: Dr. P. Pokorski University of Michigan - Dearborn Department of BiologyZuha HarithNoch keine Bewertungen
- Mitosis Vs MeiosisDocument7 pagesMitosis Vs MeiosisJD DX100% (1)
- Mitosis and Meiosis Pre-WorksheetDocument5 pagesMitosis and Meiosis Pre-WorksheetRoshae SinclairNoch keine Bewertungen
- Week 2Document8 pagesWeek 2lilyrosemary.masilangNoch keine Bewertungen
- Cell Cycle LectureDocument3 pagesCell Cycle LectureR Jay LagdaminNoch keine Bewertungen
- Science 8 Quarter 4 Shortened Module 2 CELL DIVISION Week 2Document6 pagesScience 8 Quarter 4 Shortened Module 2 CELL DIVISION Week 2Alvin OliverosNoch keine Bewertungen
- Mitosis and MeiosisDocument1 pageMitosis and Meiosisaanonymousbros.0Noch keine Bewertungen
- M2 PART 1 The Cell Cycle (READ IN ADVANCE)Document2 pagesM2 PART 1 The Cell Cycle (READ IN ADVANCE)Rizel Manlapas FernandezNoch keine Bewertungen
- MitosisDocument2 pagesMitosisGwen GwynethNoch keine Bewertungen
- Bio Discussion 3Document2 pagesBio Discussion 3api-241205310Noch keine Bewertungen
- 3 Cell Cycle, Mitosis and MeiosisDocument55 pages3 Cell Cycle, Mitosis and Meiosisapi-19641337Noch keine Bewertungen
- 1.2 Cell DivisionDocument92 pages1.2 Cell DivisionHarpreet KaurNoch keine Bewertungen
- Cell DivisionDocument17 pagesCell DivisionGenevieve GanubNoch keine Bewertungen
- Meiosis CopyDocument35 pagesMeiosis CopyAnnecalacal SabocoNoch keine Bewertungen
- CytokinesisDocument15 pagesCytokinesisMar Delos ReyesNoch keine Bewertungen
- The Cell Cycle-TRFDocument8 pagesThe Cell Cycle-TRFmaniiiiiiiiNoch keine Bewertungen
- Meiosis: Dr. Shayma'a J. Ahmed Prof. Genetic Engineerin& BiotechnologyDocument31 pagesMeiosis: Dr. Shayma'a J. Ahmed Prof. Genetic Engineerin& BiotechnologyMariam QaisNoch keine Bewertungen
- MitosisDocument24 pagesMitosisbrieNoch keine Bewertungen
- Meiosis: Chromatids (The Two Halves of A Duplicated Chromosome), As inDocument29 pagesMeiosis: Chromatids (The Two Halves of A Duplicated Chromosome), As inyamamaNoch keine Bewertungen
- BiologyDocument17 pagesBiologyMuskan ShreeNoch keine Bewertungen
- Tri-Fold Template-1Document2 pagesTri-Fold Template-1api-260947636Noch keine Bewertungen
- Mitosis & Meiosis NotesDocument6 pagesMitosis & Meiosis NotesChris_Barber09100% (1)
- Folio Kerja Kursus Biology SPMDocument29 pagesFolio Kerja Kursus Biology SPMzam_ir100% (2)
- Meiosis-Advanced 20240304 125003 0000Document25 pagesMeiosis-Advanced 20240304 125003 0000Jaja Jr. Evasco EmbarqueNoch keine Bewertungen
- Cell DivisionDocument5 pagesCell DivisionEloisa MadrilenoNoch keine Bewertungen
- Cell DivisionDocument52 pagesCell DivisionRamachandran Ram100% (1)
- Biology 3rd Edition Brooker Solutions Manual DownloadDocument11 pagesBiology 3rd Edition Brooker Solutions Manual DownloadPorfirio Dambrosio100% (21)
- Cell Cycle and Cell Division Class 11 Notes. " (Cbse)Document8 pagesCell Cycle and Cell Division Class 11 Notes. " (Cbse)madam photonNoch keine Bewertungen
- Week 5 Mitosis and MiosisDocument11 pagesWeek 5 Mitosis and MiosisEarl RénzNoch keine Bewertungen
- Biosciweek6 11Document43 pagesBiosciweek6 11Jesalyn AbrinicaNoch keine Bewertungen
- Mitosis and MeiosisDocument8 pagesMitosis and MeiosisShiela BelandresNoch keine Bewertungen
- Week 4 AssignmentDocument2 pagesWeek 4 AssignmentGood ChannelNoch keine Bewertungen
- Submission Vie4Document15 pagesSubmission Vie4Good ChannelNoch keine Bewertungen
- H03 Medical Terminology Assignment No 4Document4 pagesH03 Medical Terminology Assignment No 4Good ChannelNoch keine Bewertungen
- H02E Assignment 8Document4 pagesH02E Assignment 8Good ChannelNoch keine Bewertungen
- H06D Assignment 4Document3 pagesH06D Assignment 4Good ChannelNoch keine Bewertungen
- Question 1Document15 pagesQuestion 1Good ChannelNoch keine Bewertungen
- 0 / 2.5 Points The Medical Term For The Loss of The Ability To Eat Is Question OptionsDocument31 pages0 / 2.5 Points The Medical Term For The Loss of The Ability To Eat Is Question OptionsGood ChannelNoch keine Bewertungen
- Introduction To English Language Teaching (ENG503)Document4 pagesIntroduction To English Language Teaching (ENG503)Good ChannelNoch keine Bewertungen
- Question 14Document19 pagesQuestion 14Good ChannelNoch keine Bewertungen
- Biomes of The Forest Elephant and LifeDocument5 pagesBiomes of The Forest Elephant and LifeGood ChannelNoch keine Bewertungen
- 2.5 / 2.5 Points: SmallestDocument15 pages2.5 / 2.5 Points: SmallestGood ChannelNoch keine Bewertungen
- Assignment 8: H03 Medical TerminologyDocument5 pagesAssignment 8: H03 Medical TerminologyGood ChannelNoch keine Bewertungen
- Semester Fall 2021 Introduction To Sociology (SOC101) Assignment # 1 ScenarioDocument3 pagesSemester Fall 2021 Introduction To Sociology (SOC101) Assignment # 1 ScenarioGood ChannelNoch keine Bewertungen
- An Example of A Direct Cost Is: Question OptionsDocument15 pagesAn Example of A Direct Cost Is: Question OptionsGood ChannelNoch keine Bewertungen
- Compiled UPCAT Questions Science Ghcx2pDocument17 pagesCompiled UPCAT Questions Science Ghcx2pAdriel Pizarra100% (1)
- (Biological and Medical Physics, Biomedical Engineering) Volker Schmidt, Maria R. Belegratis (Auth.), Volker Schmidt, Maria Regina Belegratis (Eds.)-Laser Technology in Biomimetics_ Basics and ApplicaDocument281 pages(Biological and Medical Physics, Biomedical Engineering) Volker Schmidt, Maria R. Belegratis (Auth.), Volker Schmidt, Maria Regina Belegratis (Eds.)-Laser Technology in Biomimetics_ Basics and ApplicaDavy Jean DrahaNoch keine Bewertungen
- Nutrition and The Anthropology of FoodDocument22 pagesNutrition and The Anthropology of FoodSandhya MohanNoch keine Bewertungen
- 1401 - List of Companies Contains Products in Need For Validation and CompletionDocument17 pages1401 - List of Companies Contains Products in Need For Validation and CompletionNader Samir50% (2)
- 11.4 Sexual Reproduction: Essential Idea: Sexual Reproduction Involves The Development and Fusion of Haploid GametesDocument34 pages11.4 Sexual Reproduction: Essential Idea: Sexual Reproduction Involves The Development and Fusion of Haploid GametesTshegofatso100% (1)
- Pilihlah Jawaban Yang Benar Dengan Menyilang (X) Huruf A, B, Atau C!)Document4 pagesPilihlah Jawaban Yang Benar Dengan Menyilang (X) Huruf A, B, Atau C!)esty kusumaNoch keine Bewertungen
- t4w zgrTdj3Xe YcDocument10 pagest4w zgrTdj3Xe Ycvink1999Noch keine Bewertungen
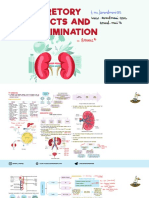
- Excretory Products and Their EliminationDocument7 pagesExcretory Products and Their EliminationKajal SinghNoch keine Bewertungen
- Glossary of Terms Used in PathologyDocument3 pagesGlossary of Terms Used in Pathologysamuel samarooNoch keine Bewertungen
- Journal of Clinical Virology: SciencedirectDocument6 pagesJournal of Clinical Virology: SciencedirectVictor VargasNoch keine Bewertungen
- Research PaperDocument4 pagesResearch Paperapi-317498189Noch keine Bewertungen
- N2o SynDocument2 pagesN2o SynethjteajNoch keine Bewertungen
- Physicochemical Properties and Antibacterial Efficacy of Ciprofloxacin - Phenazopyridine Drugs in A Binary Solid DispersionDocument5 pagesPhysicochemical Properties and Antibacterial Efficacy of Ciprofloxacin - Phenazopyridine Drugs in A Binary Solid DispersionMohammed RizkNoch keine Bewertungen
- The Functional Autonomy of MotivesDocument12 pagesThe Functional Autonomy of Motivespstrl100% (1)
- Bhs Inggris 11Document10 pagesBhs Inggris 11Kank AgoesNoch keine Bewertungen
- BSC Psychology 2017Document65 pagesBSC Psychology 2017THOMAS NJNoch keine Bewertungen
- Q4LP2 Mitosis LessonPlanDocument3 pagesQ4LP2 Mitosis LessonPlanJoshua James GitanaNoch keine Bewertungen
- Chapter 14 PP SlidesDocument58 pagesChapter 14 PP SlidesnjayrestNoch keine Bewertungen
- Eastern Visayas State University Tacloban City Riessa Jane C. Cañete Mr. Regie C. SabelaDocument12 pagesEastern Visayas State University Tacloban City Riessa Jane C. Cañete Mr. Regie C. SabelaRjane CañeteNoch keine Bewertungen
- Jurnal Pursed Lip Breathing and SaturationDocument5 pagesJurnal Pursed Lip Breathing and Saturationratna220693100% (1)
- Ecological Study of Asiaticobdella Birmanica in Lendi and Galati Stream Near Palam District Parbhani, Maharashtra IndiaDocument12 pagesEcological Study of Asiaticobdella Birmanica in Lendi and Galati Stream Near Palam District Parbhani, Maharashtra IndiaTJPRC PublicationsNoch keine Bewertungen
- RT-5 Unit 2 QPDocument7 pagesRT-5 Unit 2 QPbiotecnika_testNoch keine Bewertungen
- Encyclopedia of Infection and Immunity 1St Edition Nima Rezaei Full ChapterDocument67 pagesEncyclopedia of Infection and Immunity 1St Edition Nima Rezaei Full Chapterstephanie.stewart777100% (6)
- Class 9 MT Science Notes TissueDocument12 pagesClass 9 MT Science Notes TissueKhushi Agrawal100% (1)
- Concepts, Theories HandoutsDocument7 pagesConcepts, Theories HandoutsEdelrose LapitanNoch keine Bewertungen
- BURNS AND SCALD - Doctor SahibDocument36 pagesBURNS AND SCALD - Doctor SahibkhojalkhelNoch keine Bewertungen
- Dilla Secondary & Preparatory SchoolDocument4 pagesDilla Secondary & Preparatory SchoolGirum TesfayeNoch keine Bewertungen
- Part4 - CIE IGCSE Biology 9 RN - SampleDocument2 pagesPart4 - CIE IGCSE Biology 9 RN - SampleJun ZheNoch keine Bewertungen
- Sci8 WB IssuuDocument38 pagesSci8 WB IssuuSamanvitha RaaviNoch keine Bewertungen